Continuum Theory of Lipid Membranes
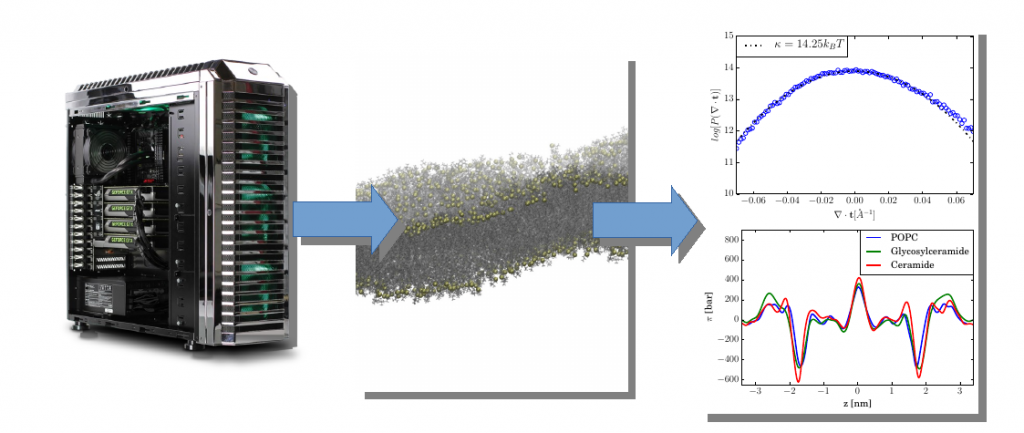
The Helfrich Theory of curvature elasticity is widely used to describe lipid membrane equilibrium shapes and deformations. In particular, it has been successfully applied to determine the shape of red blood cells. Membrane remodeling by proteins is also routinely discussed on the basis of Helfrich-type ideas. Currently, however, we do not have the theoretical model that would explain how the specific interactions of membranes influence curvature elasticity.
Using the rich information provided by Molecular Dynamics simulations in combination with our newly developed tools, we will be able to extract the specific interactions with the membrane surface.
However, curvature elasticity is but one concern when dealing with the theory of interfaces. Ions and water have also specific interactions with each other and membrane interfaces. This has lead to a widespread discontent with mean-field continuum models, such as the Poisson Boltzmann equation. We aim to incorporate the molecular structure of the interface and its solvation into new models to obtain a quantitative, coarse-grained description.
Our approach differs from ordinary coarse-grained force fields because we aim to incorporate the full information from the specific interactions into effective continuum models, which, unlike CG forcefields, contain the correct asymptotic physics.
Endosomal Escape
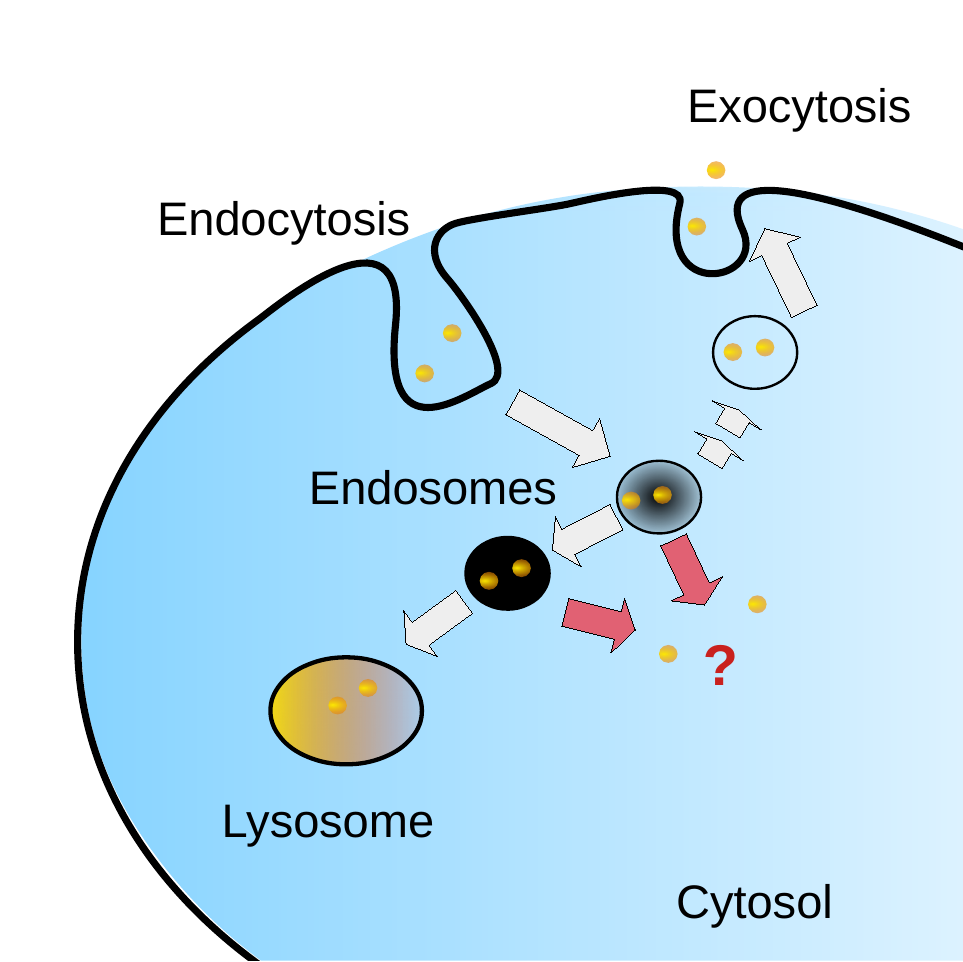
Modern complex drug delivery systems rely on an effective internalization to cells. Molecular cargo that has entered the cell via endocytosis pathways is incorporated into endosomes. If the cargo remains trapped in endosomes, the drug cannot enter the cytosol and interact with its target. Intruders, such as viruses or cellular parasites also face the same endosomal escape challenge.
In a recently published work we proposed a mechanism of cell penetration by fusion of multilamellar structures. This process may also apply for endosomal escape: Fusion of the model endosomal vesicles has been observed in experiments and they naturally contain multilamellar structures . An alternative explanation is the leakage of Bis(Monoacylglycero) Phosphate (BMP) and other endosomal model vesicles through membrane destabilization. It is, however also possible that a more indirect mechanism based on osmotic pressure or electrostatics is in play. Leakage experiments have indicated that model vesicles are selective for BMP over its isomer phosphatidylglycerol (PPG). This lipid selectivity will be used as a way of validating mechanisms of endosomal escape by cationic peptides in simulation.
Machine Learning of Interfacial Solvation

Although biophysicists traditionally describe biomembranes using continuum theories, it is natural to extend those when interfacial interactions in the presence of electrolytes need to be modeled. A well-known approach to this problem is the so-called DLVO theory, featuring the Poisson-Boltzmann equation. However, this classical mean-field theory does not include molecular structure on any level and is, therefore, unable to compute solvation mediated interactions, such as hydration forces or solvent-mediated aggregation phenomena of membranes and proteins. While there has been progress in adding some details to the Poisson-Boltzmann framework, the problems have only begun to be addressed in a systematic fashion recently.
At the same time, machine learning methods in chemistry have seen a wide interest in the past decade. Our aim is to take advantage of their abilities to describe interfacial solvation and thus pave the path to more sophisticated applications in biophysics.
Membrane Proteins

Amphipathic helical proteins, such as alpha-synuclein have long been studied for their membrane remodeling capabilities. Misfolding on the membrane is thought to influence the pathogenesis of Parkinson’s as well as Alzheimer’s disease.
Our main interest in the study of membrane proteins is the interplay of effect of lipids with intrinsic curvature, such as cardiolipin and glycosylceramides on the binding and aggregation of amphipathic helical proteins. We believe that this research will not only elucidate key factors in the genesis of important neurogenerative diseases, but will also shed some light on the mechanisms of curvature generation by specific proteins.Especially when combined with the appropriate boundary conditions. We will evaluate the damage to lipids by oxidation as well as pathological excess due to failures in biosynthesis. Our main interest in the study of membrane proteins is the interplay of effect of lipids with intrinsic curvature, such as cardiolipin and glycosylceramides on the binding and aggregation of amphipathic helical proteins.
Mitochondrial Membranes
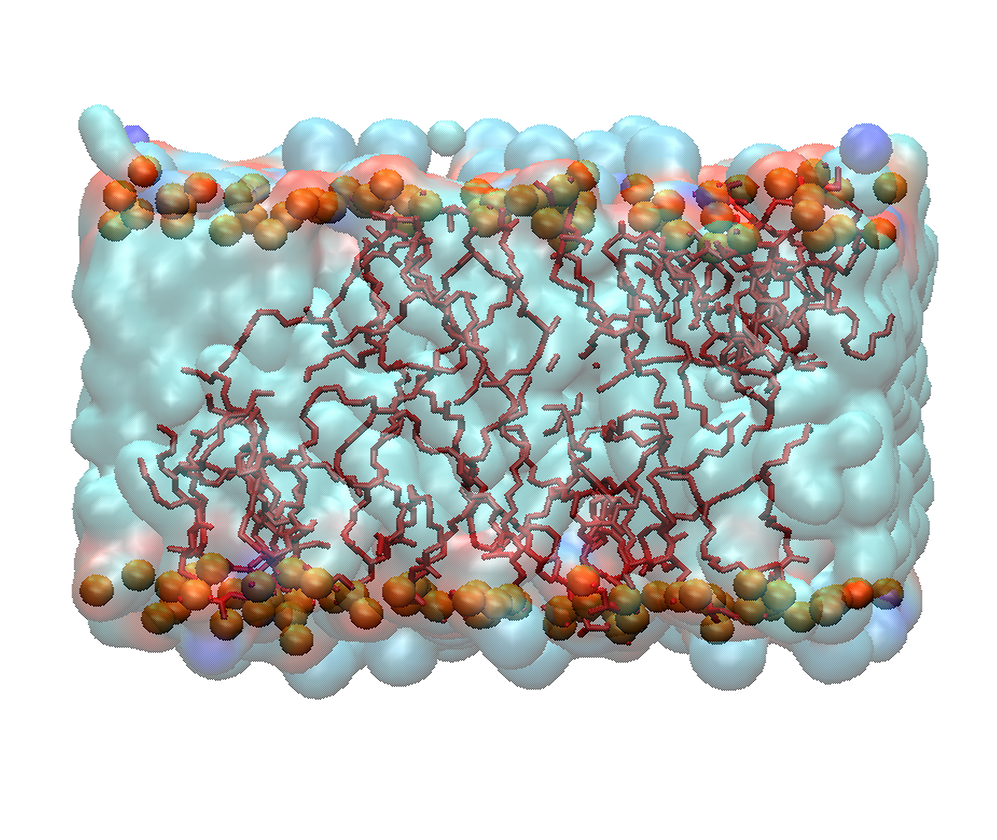
Biological membranes are highly heterogeneous in nature. Membrane physicochemical properties like fluidity, close assembly of the hydrocarbons and the membrane curvature differ from one cell to another. The structural confinement of the membrane is directly influenced by both lipid and protein composition. Membrane elastic properties regulate the cellular processes like local membrane deformation, vesicle fusion, and ion-channel function. Our main interest is to elucidate the elastic properties of mitochondrial membranes (both inner and outer).
Cardiolipin (CL), a mitochondria specific lipid, plays a crucial role in the spatial protein organization and the morphological character of the inner mitochondrial membrane (IMM). The structure of CL is quite unusual, as it has two phosphatidic acid moieties and four acyl chains. As CL has a small head group, it generates tight packing in the IMM. Any type of structural modification of CL leads to an aberrant IMM morphology and also disrupts the assembly of IMM protein complexes. Monolysocardiolipin (MLCL) is one of such modified forms that is generated in the CL remodeling pathway, leading to multisystem disease like Barth syndrome. The molecular shape of MLCL is cylindrical, whereas the shape of CL is inverted conical. In this study, we will also find out the role of oxidized CL regarding the modulation of elastic properties of the IMM.
Semiconductors in Membranes

Advanced drug delivery systems often consist of nanoparticles (NPs), such as drug-delivery liposomes. NPs can are also widely used for imaging. In this case, they are often made of metals or semiconductors with favorable fluorescence or magnetic properties and are functionalized e.g. by thiolate ligands. Ligand exchange is easy, and a wide variety of ligands is commercially available. Good control of the NP interfaces makes it possible to use these systems as an experimental testbed for endosomolytic coatings, both in cells and in model membrane experiments. It would be very attractive to also use them as detectors for changes of membrane properties.
The modification of surface states under full solvation, and even their aqueous interfaces, remain insufficiently understood and difficult to control. Therefore, we aim to functionalize semiconductor materials in order to sensitize them to their environment via their surface states. Our main interests are the response to the dielectric environment as well as to the membrane potential.